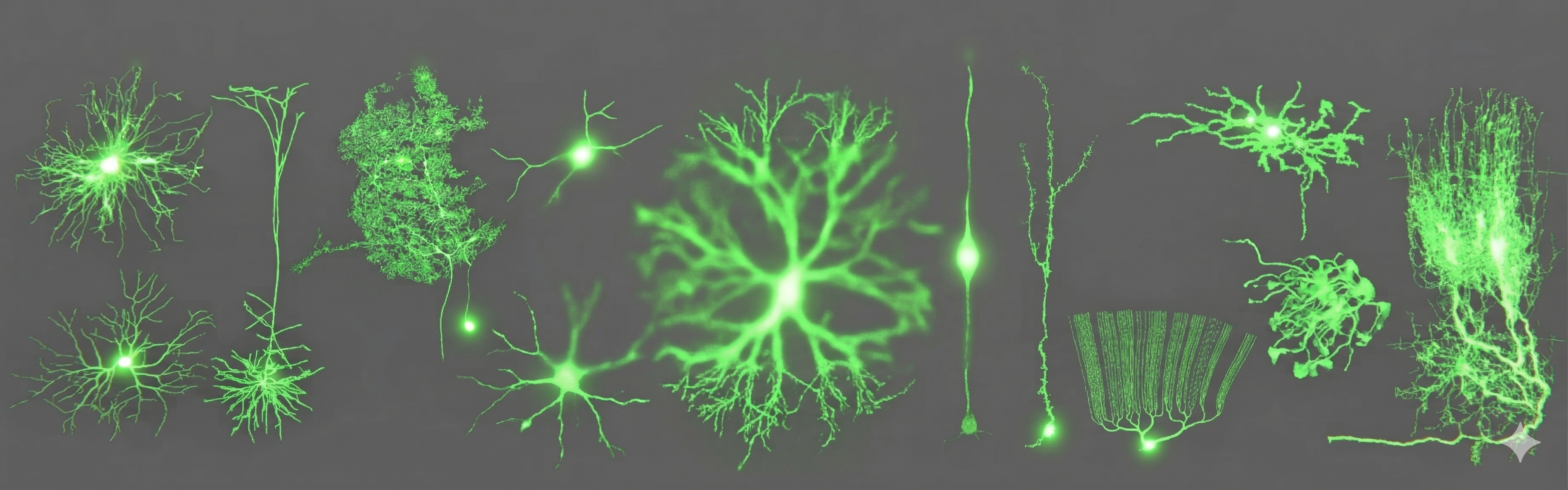
2026
Understanding how motor cortical circuits flexibly transform sensory and contextual information into behavior remains a central challenge. Whether neurons in primary vibrissal motor cortex (M1) multiplex across behaviors or are selectively engaged in context-specific actions is still unclear. To address this question, we trained mice on multiple vibrissal sensorimotor tasks, including a cue-triggered whisking-to-touch task and an air-puff–triggered licking task. Fast-spiking and regular-spiking neurons in layers 2/3 and 5 in vM1 responded robustly within ∼15 ms to air-puff stimulation. In contrast, these same neurons were only weakly modulated during goal-directed whisking-to-touch behavior. Unexpected air-puffs evoked responses in fewer neurons than expected stimuli. Trials in which stimulation elicited whisker movements produced smaller neural responses than trials without whisking. Stimulus-evoked activity in M1 was organized along a spectrum of response profiles with neurons exhibiting varying responses dynamics that cut across laminar and physiological distinctions. This organization of responses is consistent with context-dependent recruitment of M1 neurons. Together, these findings indicate that M1 activity is more closely associated with the selection of specific behavioral responses than with generalized sensory-motor encoding.
Behavioral Context Shapes Sensory Responses in Vibrissal Motor Cortex. https://doi.org/https://doi.org/10.64898/2026.04.28.720072.
Abstract Adaptive behavior relies on associating sensory cues with rewarding or aversive outcomes. In mammals, the primary sensory cortex processes stimuli and distributes information to cortical and subcortical targets. Layer 5 (L5) contains two major projection neuron classes, intratelencephalic (IT) and extratelencephalic (ET); however, their roles in associative learning remain unclear. Using transgenic mice, we identified IT and ET neurons in primary somatosensory cortex and tracked their activity with longitudinal two-photon imaging during Pavlovian conditioning with whisker stimulation. IT neurons stably encoded stimulus identity across training, whereas ET neurons showed dynamic changes that paralleled the emergence of anticipatory licking. Chemogenetic silencing of each subtype impaired learning in distinct, phase-specific ways. A reinforcement-learning model reproduced these dynamics, suggesting that IT neurons provide stable sensory representations needed to form cue-reward associations, while ET neurons encode reward expectation to refine behavior. These findings reveal complementary, cell-type-specific contributions of L5 neurons to associative learning.
Distinct roles of cortical layer 5 subtypes in associative learning. https://doi.org/10.1038/s41467-026-68307-5.
Flexible learning relies on integrating sensory and contextual information to adjust behavioral output in different environments. The anterolateral motor cortex (ALM) is a frontal area critical for action selection in rodents. We found that inputs critical to decision-making converge on the apical tuft dendrites of layer 5b pyramidal neurons in ALM. We therefore investigated the role of these dendrites in a rule-switching paradigm. Activation of dendrite-inhibiting layer 1 interneurons impaired relearning, without affecting previously learned behavior. This inhibition profoundly suppressed global calcium activity in dendritic shafts but not local transients in spines, while additionally reducing burst firing. Moreover, excitatory synaptic inputs to tuft dendrites exhibited rule-dependent clustering. We conclude that dendritic calcium signaling is a key computational component of flexible learning. , Editor’s summary Flexible adaptive behavior is a hallmark of intelligent systems and requires continuous integration of sensory information and the ability to optimize motor plans in a context-dependent manner. Maristany de las Casas et al . investigated the neuronal mechanisms underlying flexible learning in the mouse anterolateral motor cortex. They used a rule-switching exercise from complex to simple and back to complex tasks. Nonlinear dendritic events were essential for relearning in these tasks. Active dendritic integration is thus a potential mechanism enabling the brain to solve the cognitive challenges posed by adaptive behavior. —Peter Stern , INTRODUCTION The ability to flexibly adapt behavior in changing environments is a hallmark of intelligence, yet the neural mechanisms underlying this capability remain poorly understood. In the mammalian brain, pyramidal neurons (the primary excitatory cells of the cerebral cortex) possess elaborate dendritic trees that extend far from the cell body. These dendrites have two main compartments, corresponding to the main dendritic arborizations (the basal and apical tuft dendrites). The basal compartment is thought to receive feature-specific information (e.g., sensory features in sensory cortex), whereas the apical tuft compartment receives contextual information (e.g., information from an internal source). Although neurons are often modeled as simple integrators that sum their inputs, emerging evidence, including this study, suggests that dendrites can perform complex computations through active electrical properties, potentially enabling sophisticated learning capabilities. RATIONALE We hypothesized that dendritic computations in frontal motor areas contribute to flexible behavioral adaptation. Apical dendrites of layer 5 pyramidal neurons in the anterolateral motor cortex (ALM) in mice receive convergent inputs from sensory and motor-planning regions. These dendrites can generate calcium-dependent electrical events that may serve as a substrate for learning. To test this hypothesis, we developed a behavioral paradigm where mice must flexibly switch between different stimulus-response rules, allowing us to dissociate task execution from the cognitive demands of relearning. We focused on a specific population of inhibitory neurons [NDNF (neuron-derived neurotrophic factor) interneurons] that selectively target the apical dendrites, providing the means to manipulate dendritic activity. RESULTS Using a combination of behavioral testing, optical recordings, and targeted neural manipulations, we discovered that calcium activity in the apical tuft dendrite is selectively required for relearning complex, but not simple, behavioral rules. When mice performed our rule-switching task, activating dendrite-inhibiting NDNF interneurons impaired their ability to relearn the complex rule after exposure to the simpler one, while leaving performance of already-learned behaviors unchanged. Two-photon (2P) calcium imaging and electrophysiological recordings revealed that this manipulation abolished global calcium events throughout the dendritic tree while preserving local synaptic activity and only moderately affecting somatic action potentials. Imaging of synaptic inputs to dendrites revealed that they organize into functional clusters during complex rule performance—a spatial arrangement that dissolved during simple rule execution. Notably, the NDNF interneurons themselves reduced their activity specifically when mice made errors during rule switching (and initial learning), suggesting that they gate dendritic plasticity. CONCLUSION Our findings reveal that calcium signaling in dendritic tufts serves as a critical computational resource for flexible learning. This work establishes that cortical circuits can selectively engage active dendritic computation on the basis of cognitive demands, with inhibitory control providing dynamic gating of these processes during learning. These mechanisms may explain how the brain maintains stable behaviors while retaining the capacity for rapid adaptation, with implications for understanding cognitive flexibility disorders and developing therapeutic interventions. More broadly, our results suggest that the elaborate dendritic trees of pyramidal neurons are not merely passive conduits but active computational units essential for flexible behavior. Flexible learning is dependent on apical dendritic calcium activity. Mice were trained to transition between a complex task (rule A) and a simple task (rule B). Activation of layer 1 (L1) NDNF interneurons blocked relearning of complex, but not simpler, tasks through the suppression of calcium throughout the apical tuft but not locally in spines. Functional clustering of synapses was associated with task complexity.
Tuft dendrites in frontal motor cortex enable flexible learning. https://doi.org/10.1126/science.adx4358.
Could a long-overlooked cortical layer hold the key to attention? Essential for our perception and everyday experience, attention is thought to depend on precisely controlled feedback loops between cortical layer 5 and the higher-order thalamus. However, despite their critical importance, the circuits that precisely control these ‘‘attention loops’’ remain poorly understood. Interestingly, recent advances revealed that the deepest cortical neurons, layer 6b (L6b), may be a powerful yet previously unrecognized conductor of these feedback loops. L6b is positioned at the intersection of top-down cortical drive and convergent neuromodulation and its projections appear uniquely equipped to precisely control these loops. Here, we propose the L6b attention theory (LAT), a circuit-based model that complements and extends existing theories of attention. LAT suggests that this overlooked layer is a missing link in our understanding of attention and cognition, offering new insights into neuropsychiatric disorders and how we understand our mental state.
The layer 6b theory of attention. https://doi.org/10.1016/j.neuron.2025.11.024.
2025
A key function of the brain is to move the body through a rich, complex environment. When rodents engage with their environment, they move their whiskers as they extract tactile information. Even though the study of whisking has a long history, the details of individual whisker movements bilaterally, of nose movement, of stereotypy and variability in an active whisking to touch behavior are unknown. Here we trained five head-fixed male and 11 female mice in a go-cue task to move a whisker to touch a sensor. Our analysis of orofacial movements shows that mice specifically control movement of the whisker they use to touch and that as they move their whiskers, they move their nose and apply forces on the head post in a manner that reflects the behavioral epoch, i.e., whether go-cue triggered movement had begun or a whisker was touching the sensor. Importantly, mice control the setpoint, amplitude, and frequency of movement of whiskers bilaterally and individually. Additionally, even though mice achieved the goal of the task—to touch the sensor within 2 s—how they coordinated movement of the nose and forces on head post with movement of individual whiskers was stereotyped and related to the distance they needed to move a whisker to touch the sensor. Our work shows how stereotyped mouse behavior can be, and it emphasizes both the level of fine motor control mice can exert over individual whiskers and the extent of facial movements in a goal-directed whisking-to-touch task.
Orofacial Movements: Individuality and Stereotypy When Mice Move a Single Whisker to Touch. https://doi.org/10.1523/JNEUROSCI.0712-25.2025.
Head fixation of rodents is a widely utilized and important technique that enables laboratories to measure brain activity during behavior, but head fixation can increase stress which affects both behavior and underlying brain activity, as well as animal welfare. It is therefore critical to keep stress levels low, yet it is particularly challenging to assess stress in immobilized, head-fixed rodents. Conventional stress evaluation methods, such as blood corticosterone analysis, are labor-intensive and conducted post hoc, and In situ approaches require experienced personnel and constant vigilance during experiments. In this study, we present MouseCare, an automated software solution for immediate stress detection by real-time facial feature video analysis in head-fixed mice. MouseCare performs on par with or better than conventional stress measures, and is significantly less labor-intensive. This approach enables objective measures of stress that are needed to determine when an experiment can commence, but also when it should be stopped. We conclude that MouseCare offers a cost-effective and easily implementable strategy to manage stress levels in rodents that can increase data quality and improve animal welfare.
Automated facial feature evaluation system to prevent stress of head fixed mice. https://doi.org/10.1371/journal.pone.0322530.
Acute Optogenetic Stimulation of Serotonin Neurons Reduces Cell Proliferation in the Dentate Gyrus of Mice. https://doi.org/10.1021/acschemneuro.4c00771.
Feeding-induced olfactory cortex suppression reduces satiation. https://doi.org/10.1016/j.neuron.2025.07.020.
Artificial neural networks are becoming more advanced and human-like in detail and behavior. The notion that machines mimicking human brain computations might be conscious has recently caused growing unease. Here, we explored a common computational functionalist view, which holds that consciousness emerges when the right computations occur—whether in a machine or a biological brain. To test this view, we simulated a simple computation in an artificial subject’s “brain” and recorded each neuron’s activity when the subject was presented with a visual stimulus. We then replayed these recorded signals back into the same neurons, degrading the computation by effectively eliminating all alternative activity patterns that otherwise might have occurred (i.e., the counterfactuals). We identified a special case in which the replay did nothing to the subject’s ongoing brain activity—allowing it to evolve naturally in response to a stimulus—but still degraded the computation by erasing the counterfactuals. This paradoxical outcome points to a disconnect between ongoing neural activity and the underlying computational structure, which challenges the notion that consciousness arises from computation in artificial or biological brains.
Does neural computation feel like something?. https://doi.org/10.3389/fnins.2025.1511972.
Bimodal nonlinear dendrites in PV+ basket cells drive distinct memory-related oscillations. https://doi.org/10.1016/j.isci.2025.113699.
Contemporary models of perceptual awareness lack tractable neurobiological constraints. Inspired by recent cellular recordings in a mouse model of tactile threshold detection, we constructed a biophysical model of perceptual awareness that incorporated essential features of thalamocortical anatomy and cellular physiology. Our model reproduced, and mechanistically explains, the key in vivo neural and behavioural signatures of perceptual awareness in the mouse model, as well as the response to a set of causal perturbations. We generalised the same model (with identical parameters) to a more complex task – visual rivalry – and found that the same thalamic-mediated mechanism of perceptual awareness determined perceptual dominance. This led to the generation of a set of novel, and directly testable, electrophysiological predictions. Analyses of the model based on dynamical systems theory show that perceptual awareness in simulations of both threshold detection and visual rivalry arises from the emergent systems-level dynamics of thalamocortical loops.
A burst-dependent thalamocortical substrate for perceptual awareness. https://doi.org/10.1371/journal.pcbi.1012951.
2024
How is conscious experience related to material brain processes? A variety of theories aiming to answer this age-old question have emerged from the recent surge in consciousness research, and some are now hotly debated. Although most researchers have so far focused on the development and validation of their preferred theory in relative isolation, this article, written by a group of scientists representing different theories, takes an alternative approach. Noting that various theories often try to explain different aspects or mechanistic levels of consciousness, we argue that the theories do not necessarily contradict each other. Instead, several of them may converge on fundamental neuronal mechanisms and be partly compatible and complementary, so that multiple theories can simultaneously contribute to our understanding. Here, we consider unifying, integration-oriented approaches that have so far been largely neglected, seeking to combine valuable elements from various theories.
An integrative, multiscale view on neural theories of consciousness. https://doi.org/10.1016/j.neuron.2024.02.004.
The deepest layer of the cortex (layer 6b [L6b]) contains relatively few neurons, but it is the only cortical layer responsive to the potent wake-promoting neuropeptide orexin/hypocretin. Can these few neurons significantly influence brain state? Here, we show that L6b-photoactivation causes a surprisingly robust enhancement of attention-associated high-gamma oscillations and population spiking while abolishing slow waves in sleep-deprived mice. To explain this powerful impact on brain state, we investigated L6b’s synaptic output using optogenetics, electrophysiology, and monoCaTChR ex vivo. We found powerful output in the higher-order thalamus and apical dendrites of L5 pyramidal neurons, via L1a and L5a, as well as in superior colliculus and L6 interneurons. L6b subpopulations with distinct morphologies and short- and long-term plasticities project to these diverse targets. The L1a-targeting subpopulation triggered powerful NMDA-receptor-dependent spikes that elicited burst firing in L5. We conclude that orexin/hypocretin-activated cortical neurons form a multifaceted, fine-tuned circuit for the sustained control of the higher-order thalamocortical system.
Layer 6b controls brain state via apical dendrites and the higher-order thalamocortical system. https://doi.org/10.1016/j.neuron.2023.11.021.
Abstract In recent years, brain research has indisputably entered a new epoch, driven by substantial methodological advances and digitally enabled data integration and modelling at multiple scales—from molecules to the whole brain. Major advances are emerging at the intersection of neuroscience with technology and computing. This new science of the brain combines high-quality research, data integration across multiple scales, a new culture of multidisciplinary large-scale collaboration, and translation into applications. As pioneered in Europe’s Human Brain Project (HBP), a systematic approach will be essential for meeting the coming decade’s pressing medical and technological challenges. The aims of this paper are to: develop a concept for the coming decade of digital brain research, discuss this new concept with the research community at large, identify points of convergence, and derive therefrom scientific common goals; provide a scientific framework for the current and future development of EBRAINS, a research infrastructure resulting from the HBP’s work; inform and engage stakeholders, funding organisations and research institutions regarding future digital brain research; identify and address the transformational potential of comprehensive brain models for artificial intelligence, including machine learning and deep learning; outline a collaborative approach that integrates reflection, dialogues, and societal engagement on ethical and societal opportunities and challenges as part of future neuroscience research.
The coming decade of digital brain research: A vision for neuroscience at the intersection of technology and computing. https://doi.org/10.1162/imag_a_00137.
ABSTRACT Neurons in primary visual cortex are driven by feedforward visual inputs and top-down contextual inputs. The nature of this contextual information is difficult to study, as responses to feedforward and top-down inputs overlap in time and are difficult to disentangle experimentally. To address this issue, we measured responses to natural images and partially occluded versions of these images in the visual cortex of mice. Assessing neuronal responses before and after familiarizing mice with the non-occluded images allowed us to study experience-dependent and stimulus-specific contextual responses in pyramidal cells (PyCs) in cortical layers 2/3 and 5 in the absence of feedforward input. Surprisingly, in the same retinotopic region of cortex, we found that separate populations of PyCs in layer 2/3 responded to occluded and non-occluded images. Responses of PyCs selective for occluded images were strengthened upon familiarization and decoding analysis revealed they contained image-specific information, suggesting that they signaled the absence of predicted visual stimuli. Responses of PyCs selective for non-occluded scenes were weaker for familiarized images but stronger for unfamiliar images, suggesting that these neurons signaled the presence of unpredicted visual stimuli. Layer 5 also contained PyCs preferring either feedforward or contextual inputs, but their responses were more complex and strengthening of responses to occluded images required task engagement. The results show that visual experience decreases the activity of neurons responding to known feedforward inputs but increases the activity of neurons responding to contextual inputs tied to expected stimuli.
Experience-dependent predictions of feedforward and contextual information in mouse visual cortex. https://doi.org/10.1101/2024.06.10.598181.
Theories of attention and learning have hypothesized a central role for high-frequency bursting in cognitive functions, but experimental reports of burst-mediated representations in vivo have been limited. Here we used a novel demultiplexing approach by considering a conjunctive burst code. We studied this code in vivo while animals learned to report direct electrical stimulation of the somatosensory cortex and found two acquired yet independent representations. One code, the event rate, showed a sparse and succint stiumulus representation and a small modulation upon detection errors. The other code, the burst fraction, correlated more globally with stimulation and more promptly responded to detection errors. Bursting modulation was potent and its time course evolved, even in cells that were considered unresponsive based on the firing rate. During the later stages of training, this modulation in bursting happened earlier, gradually aligning temporally with the representation in event rate. The alignment of bursting and event rate modulation sharpened the firing rate response, and was strongly associated behavioral accuracy. Thus a fine-grained separation of spike timing patterns reveals two signals that accompany stimulus representations: an error signal that can be essential to guide learning and a sharpening signal that could implement attention mechanisms.
Fast burst fraction transients convey information independent of the firing rate. https://doi.org/10.7554/eLife.98290.1.
Summary Not all decisions are created equal; factors such as the difficulties or associated costs affect the time spent to make decisions. This is variously interpreted as speed/accuracy, fast/slow, or impulsivity/deliberateness tradeoffs according to different models of behaviour 1–5 . Regardless, it is generally assumed that decision latency reflects the neural mechanisms underlying behavioural strategy and cognitive investment. However, such investigations have been difficult in mice which are consistently impulsive. Here, we show that manipulating cost, using a novel floating-platform paradigm, overcomes the natural impulsivity of mice, more closely matching human behaviour. Furthermore, this approach allowed us simultaneously to measure the flow of activity from medial to lateral frontal cortex (MFC→LFC) and record sequences of single neuron activity with 2-photon imaging. Surprisingly, MFC display a different mode of operation, with high vulnerability to optical inhibition compared to LFC. Furthermore, the balance in choice coding at the beginning of sequences in MFC correlated with trial history and behavioural strategy. We found that for optimal performance, slow sequences in MFC showed declining numbers of active neurons whereas the opposite was true in LFC. Our results suggest that while LFC acts as an integrative motor threshold, MFC plays a larger cognitive role in the selection and timing of decisions than previously thought. Our study offers a methodological and mechanistic framework in mouse frontal cortex to understand the neural basis of voluntary decision making.
The neural mechanisms of fast versus slow decision-making. https://doi.org/10.1101/2024.08.22.608577.
Abstract Hippocampal oscillations regulate distinct memory stages and behavioral states, yet the mechanisms by which interneurons contribute to these rhythms remain elusive. Here, we establish a novel link between the bimodal nonlinear dendritic processing of PV+ fast-spiking basket cells (FSBCs) and hippocampal oscillations. Our model predicts that FSBCs can flexibly adjust their activity without changes in synaptic input quantity, leveraging their supralinear and sublinear dendritic branches to selectively shape network dynamics. Supralinear dendritic activation enhances high-frequency oscillations, while sublinear dendritic activation amplifies slow oscillatory power, dynamically modulating the excitation/inhibition (E/I) balance. These findings reveal a previously unrecognized role of FSBC dendritic integration in memory-related oscillatory dynamics, offering new insights into the cellular mechanisms governing hippocampal rhythm generation. Significance Statement Hippocampal oscillations play a crucial role in memory processes, yet how interneurons dynamically regulate these rhythms remains unresolved. Our study reveals that fast-spiking basket cells (FSBCs) exploit bimodal nonlinear dendritic processing to selectively shape slow and fast oscillatory components, independently of synaptic input quantity. This energy-efficient mechanism allows FSBCs to fine-tune the excitation-inhibition balance, modulating network dynamics essential for learning and consolidation. By challenging the conventional view of interneuron function, our findings redefine how inhibitory circuits contribute to hippocampal rhythm generation, providing new insights into the cellular basis of memory-related oscillations.
Bimodal dendritic processing in basket cells drives distinct memory-related oscillations. https://doi.org/10.1101/2024.09.11.612262.
High-speed volumetric imaging is crucial for observing fast and distributed processes such as neuronal activity. Multiphoton microscopy helps to mitigate scattering effects inside tissue, but the standard raster scanning approach limits achievable volume rates. Random access point scanning can lead to a considerable speed-up by sampling only pre-selected locations, but existing techniques based on acousto-optic deflectors are still limited to a point rate of up to . This limits the number of parallel targets at the high acquisition rates necessary, for example, in voltage imaging or imaging of fast synaptic events. Here, we introduce SPARCLS, a method for 3D random access point scanning at up to 340 kHz using a single 1D phase modulator. We show the potential of this method by imaging synaptic events with fluorescent glutamate sensors in mammalian organotypic slices as well as in zebrafish larvae.
High-speed three-dimensional random access scanning with a linear SLM. https://doi.org/10.1364/OPTICA.536853.
Accurate assessment of post-stroke deficits is crucial in translational research. Recent advances in machine learning offer precise quantification of rodent motor behavior post-stroke, yet detecting lesion-specific upper extremity deficits remains unclear. Employing proximal middle cerebral artery occlusion (MCAO) and cortical photothrombosis (PT) in mice, we assessed post-stroke impairments via the Staircase test. Lesion locations were identified using 7 T-MRI. Machine learning was applied to reconstruct forepaw kinematic trajectories and feature analysis was achieved with MouseReach, a new data-processing toolbox. Lesion reconstructions pinpointed ischemic centers in the striatum (MCAO) and sensorimotor cortex (PT). Pellet retrieval alterations were observed, but were unrelated to overall stroke volume. Instead, forepaw slips and relative reaching success correlated with increasing cortical lesion size in both models. Striatal lesion size after MCAO was associated with prolonged reach durations that occurred with delayed symptom onset. Further analysis on the impact of selective serotonin reuptake inhibitors in the PT model revealed no clear treatment effects but replicated strong effect sizes of slips for post-stroke deficit detection. In summary, refined movement analysis unveiled specific deficits in two widely-used mouse stroke models, emphasizing the value of deep behavioral profiling in preclinical stroke research to enhance model validity for clinical translation.
Refined movement analysis in the staircase test reveals differential motor deficits in mouse models of stroke. https://doi.org/10.1177/0271678x241254718.
Pyramidal neurons have a pivotal role in the cognitive capabilities of neocortex. Though they have been pre dominantly modeled as integrate-and-fire point processors, many of them have another point of input integration in their apical dendrites that is central to mechanisms endowing them with the sensitivity to context that un derlies basic cognitive capabilities. Here we review evidence implicating impairments of those mechanisms in three major neurodevelopmental disabilities, fragile X, Down syndrome, and fetal alcohol spectrum disorders. Multiple dysfunctions of the mechanisms by which pyramidal cells are sensitive to context are found to be implicated in all three syndromes. Further deciphering of these cellular mechanisms would lead to the under standing of and therapies for learning disabilities beyond any that are currently available.
Dysfunctions of cellular context-sensitivity in neurodevelopmental learning disabilities. https://doi.org/10.1016/j.neubiorev.2024.105688.
The hippocampus is crucial for acquiring and retrieving episodic and contextual memories. In previous studies, the inactivation of dentate gyrus (DG) neurons by chemogenetic- and optogenetic-mediated hyperpolarization led to opposing conclusions about DG’s role in memory retrieval. One study used Designer Receptors Exclusively Activated by Designer Drugs (DREADD)-mediated clozapine N-oxide (CNO)-induced hyperpolarization and reported that the previously formed memory was erased, thus concluding that denate gyrus is needed for memory maintenance. The other study used optogenetic with halorhodopsin induced hyperpolarization and reported and dentate gyrus is needed for memory retrieval. We hypothesized that this apparent discrepancy could be due to the length of hyperpolarization in previous studies; minutes by optogenetics and several hours by DREADD/CNO. Since hyperpolarization interferes with anterograde and retrograde neuronal signaling, it is possible that the memory engram in the dentate gyrus and the entorhinal to hippocampus trisynaptic circuit was erased by long-term, but not with short-term hyperpolarization. We developed and applied an advanced chemogenetic technology to selectively silence synaptic output by blocking neurotransmitter release without hyperpolarizing DG neurons to explore this apparent discrepancy. We performed in vivo electrophysiology during trace eyeblink in a rabbit model of associative learning. Our work shows that the DG output is required for memory retrieval. Based on previous and recent findings, we propose that the actively functional anterograde and retrograde neuronal signaling is necessary to preserve synaptic memory engrams along the entorhinal cortex to the hippocampal trisynaptic circuit.
Dentate gyrus is needed for memory retrieval. https://doi.org/10.1038/s41380-024-02546-0.
2023
Contemporary models of perceptual awareness lack tractable neurobiological constraints. Inspired by recent cellular recordings in a mouse model of tactile threshold detection, we constructed a biophysical model of perceptual awareness that incorporated essential features of thalamocortical anatomy and cellular physiology. Our model reproduced, and mechanistically explains, the key in vivo neural and behavioural signatures of perceptual awareness in the mouse model, as well as the response to a set of causal perturbations. We generalised the same model (with identical parameters) to a more complex task – visual rivalry – and found that the same thalamic-mediated mechanism of perceptual awareness determined perceptual dominance. This led to the generation of a set of novel, and directly testable, electrophysiological predictions. Analyses of the model based on dynamical systems theory show that perceptual awareness in simulations of both threshold detection and visual rivalry arises from the emergent systems-level dynamics of thalamocortical loops.
A Burst-dependent Thalamocortical Substrate for Perceptual Awareness. https://doi.org/10.1101/2023.07.13.548934.
How do new ideas come about? The central hypothesis presented here states that insights might happen during mental navigation and correspond to rapid plasticity at the cellular level. We highlight the differences between neocortical and hippocampal mechanisms of insight. We argue that the suddenness of insight can be related to the sudden emergence of place fields in the hippocampus. According to our hypothesis, insights are supported by a state of mind-wandering that can be tied to the process of combining knowledge pieces during sharp-wave ripples (SWRs). Our framework connects the dots between research on creativity, mental navigation, and specific synaptic plasticity mechanisms in the hippocampus.
Mental navigation and the neural mechanisms of insight. https://doi.org/10.1016/j.tins.2022.11.002.
Interactions with large language models (LLMs) have led to the suggestion that these models may soon be conscious. From the perspective of neuroscience, this position is difficult to defend. For one, the inputs to LLMs lack the embodied, embedded information content characteristic of our sensory contact with the world around us. Secondly, the architectures of present-day artificial intelligence algorithms are missing key features of the thalamocortical system that have been linked to conscious awareness in mammals. Finally, the evolutionary and developmental trajectories that led to the emergence of living conscious organisms arguably have no parallels in artificial systems as envisioned today. The existence of living organisms depends on their actions and their survival is intricately linked to multi-level cellular, inter-cellular, and organismal processes culminating in agency and consciousness.
The feasibility of artificial consciousness through the lens of neuroscience. https://doi.org/10.1016/j.tins.2023.09.009.
Abstract Neocortical layer 1 has been proposed to be at the center for top-down and bottom-up integration. It is a locus for interactions between long-range inputs, layer 1 interneurons, and apical tuft dendrites of pyramidal neurons. While input to layer 1 has been studied intensively, the level and effect of input to this layer has still not been completely characterized. Here we examined the input to layer 1 of mouse somatosensory cortex with retrograde tracing and optogenetics. Our assays reveal that local input to layer 1 is predominantly from layers 2/3 and 5 pyramidal neurons and interneurons, and that subtypes of local layers 5 and 6b neurons project to layer 1 with different probabilities. Long-range input from sensory-motor cortices to layer 1 of somatosensory cortex arose predominantly from layers 2/3 neurons. Our optogenetic experiments showed that intra-telencephalic layer 5 pyramidal neurons drive layer 1 interneurons but have no effect locally on layer 5 apical tuft dendrites. Dual retrograde tracing revealed that a fraction of local and long-range neurons was both presynaptic to layer 5 neurons and projected to layer 1. Our work highlights the prominent role of local inputs to layer 1 and shows the potential for complex interactions between long-range and local inputs, which are both in position to modify the output of somatosensory cortex.
Layer 1 of somatosensory cortex: an important site for input to a tiny cortical compartment. https://doi.org/10.1093/cercor/bhad371.
The organization of fear memory involves the participation of multiple brain regions. However, it is largely unknown how fear memory is formed, which circuit pathways are used for “printing” memory engrams across brain regions, and the role of identified brain circuits in memory retrieval. With advanced genetic methods, we combinatorially blocked presynaptic output and manipulated N-methyl-D-aspartate receptor (NMDAR) in the basolateral amygdala (BLA) and medial prefrontal cortex (mPFC) before and after cued fear conditioning. Further, we tagged fear-activated neurons during associative learning for optogenetic memory recall. We found that presynaptic mPFC and postsynaptic BLA NMDARs are required for fear memory formation, but not expression. Our results provide strong evidence that NMDAR-dependent synaptic plasticity drives multi-trace systems consolidation for the sequential printing of fear memory engrams from BLA to mPFC and, subsequently, to the other regions, for flexible memory retrieval.
Pre- and postsynaptic N-methyl-D-aspartate receptors are required for sequential printing of fear memory engrams. https://doi.org/10.1016/j.isci.2023.108050.
Understanding the neurobiological mechanisms underlying consciousness remains a significant challenge. Recent evidence suggests that the coupling between distal–apical and basal–somatic dendrites in thick-tufted layer 5 pyramidal neurons (L5 PN ), regulated by the nonspecific-projecting thalamus, is crucial for consciousness. Yet, it is uncertain whether this thalamocortical mechanism can support emergent signatures of consciousness, such as integrated information. To address this question, we constructed a biophysical network of dual-compartment thick-tufted L5 PN , with dendrosomatic coupling controlled by thalamic inputs. Our findings demonstrate that integrated information is maximized when nonspecific thalamic inputs drive the system into a regime of time-varying synchronous bursting. Here, the system exhibits variable spiking dynamics with broad pairwise correlations, supporting the enhanced integrated information. Further, the observed peak in integrated information aligns with criticality signatures and empirically observed layer 5 pyramidal bursting rates. These results suggest that the thalamocortical core of the mammalian brain may be evolutionarily configured to optimize effective information processing, providing a potential neuronal mechanism that integrates microscale theories with macroscale signatures of consciousness.
A thalamocortical substrate for integrated information via critical synchronous bursting. https://doi.org/10.1073/pnas.2308670120.
2022
Abstract The lipid phosphatase PTEN (phosphatase and tensin homologue on chromosome 10) is a key tumour suppressor gene and an important regulator of neuronal signalling. PTEN mutations have been identified in patients with autism spectrum disorders, characterized by macrocephaly, impaired social interactions and communication, repetitive behaviour, intellectual disability, and epilepsy. PTEN enzymatic activity is regulated by a cluster of phosphorylation sites at the C-terminus of the protein. Here, we focused on the role of PTEN T366 phosphorylation and generated a knock-in mouse line in which Pten T366 was substituted with alanine (PtenT366A/T366A). We identify that phosphorylation of PTEN at T366 controls neuron size and connectivity of brain circuits involved in sensory processing. We show in behavioural tests that PtenT366/T366A mice exhibit cognitive deficits and selective sensory impairments, with significant differences in male individuals. We identify restricted cellular overgrowth of cortical neurons in PtenT366A/T366A brains, linked to increases in both dendritic arborization and soma size. In a combinatorial approach of anterograde and retrograde monosynaptic tracing using rabies virus, we characterize differences in connectivity to the primary somatosensory cortex of PtenT366A/T366A brains, with imbalances in long-range cortico-cortical input to neurons. We conclude that phosphorylation of PTEN at T366 controls neuron size and connectivity of brain circuits involved in sensory processing and propose that PTEN T366 signalling may account for a subset of autism-related functions of PTEN.
The impact of phosphorylated PTEN at threonine 366 on cortical connectivity and behaviour. https://doi.org/10.1093/brain/awac188.
To strengthen the translational value of the Chronic Social Defeat male mouse model in stress resilience research, we are using Magnetic Resonance Imaging and Diffusion Tensor Imaging to understand the underlying brain connectivity and anatomy of the different behavioural phenotypic readouts (stress resilient, stress susceptible, and non-learners) observed within a single defeated group as identified using the Modified Social Interaction Test.
Study protocol: Brain region-specific signatures in connectivity and anatomy between stress resilient and susceptible mice. https://doi.org/10.17605/OSF.IO/MDTH4.
This article presents the argument that, while understanding the brain will require a multi-level approach, there is nevertheless something fundamental about understanding the components of the brain. I argue here that the standard description of neurons is not merely too simplistic, but also misses the true nature of how they operate at the computational level. In particular, the humble point neuron, devoid of dendrites with their powerful computational properties, prevents conceptual progress at higher levels of understanding.
Are Dendrites Conceptually Useful?. https://doi.org/10.1016/j.neuroscience.2022.03.008.
Half a century since their discovery by Llinás and colleagues, dendritic spikes have been observed in various neurons in different brain regions, from the neocortex and cerebellum to the basal ganglia. Dendrites exhibit a terrifically diverse but stereotypical repertoire of spikes, sometimes specific to subregions of the dendrite. Despite their prevalence, we only have a glimpse into their role in the behaving animal. This article aims to survey the full range of dendritic spikes found in excitatory and inhibitory neurons, compare them in vivo versus in vitro, and discuss new studies describing dendritic spikes in the human cortex. We focus on neocortical and hippocampal neurons and present a roadmap to identify and understand the broader role of dendritic spikes in single-cell computation.
The Guide to Dendritic Spikes of the Mammalian Cortex In Vitro and In Vivo. https://doi.org/10.1016/j.neuroscience.2022.02.009.
In mammalian neocortex, learning triggers the formation and turnover of new postsynaptic spines on pyramidal cell dendrites. However, the biological principles of spine reorganization during learning remain elusive because the identity of their presynaptic neuronal partners is unknown. Here, we show that two presynaptic neural circuits supervise distinct programs of spine dynamics to execute learning. We imaged spine dynamics in motor cortex during learning and performed post hoc identification of their afferent presynaptic neurons. New spines that appeared during learning formed small transient contacts with corticocortical neurons that were eliminated on skill acquisition. In contrast, persistent spines with axons from thalamic neurons were formed and enlarged. These results suggest that pyramidal cell dendrites in motor cortex use a neural circuit division of labor during skill learning, with dynamic teaching contacts from top-down intracortical axons followed by synaptic memory formation driven by thalamic axons. Dual spine supervision may govern diverse skill learning in the neocortex. , Corticocortical and thalamocortical neural circuits presynaptically determine postsynaptic spine dynamics during learning.
Presynaptic supervision of cortical spine dynamics in motor learning. https://doi.org/10.1126/sciadv.abm0531.
Abstract Navigation through complex environments requires motor planning, motor preparation, and the coordination between multiple sensory–motor modalities. For example, the stepping motion when we walk is coordinated with motion of the torso, arms, head, and eyes. In rodents, movement of the animal through the environment is coordinated with whisking. Even head-fixed mice navigating a plus maze position their whiskers asymmetrically with the bilateral asymmetry signifying the upcoming turn direction. Here we report that, in addition to moving their whiskers, on every trial mice also move their eyes conjugately in the direction of the upcoming turn. Not only do mice move their eyes, but they coordinate saccadic eye movement with the asymmetric positioning of the whiskers. Our analysis shows that asymmetric positioning of whiskers predicted the turn direction that mice will make at an earlier stage than eye movement. Consistent with these results, our observations also revealed that whisker asymmetry increases before saccadic eye movement. Importantly, this work shows that when rodents plan for active behavior, their motor plans can involve both eye and whisker movement. We conclude that, when mice are engaged in and moving through complex real-world environments, their behavioral state can be read out in the movement of both their whiskers and eyes.
Coordination between Eye Movement and Whisking in Head-Fixed Mice Navigating a Plus Maze. https://doi.org/10.1523/ENEURO.0089-22.2022.
Rapid advances in neuroscience have provided remarkable breakthroughs in understanding the brain on many fronts. Although promising, the role of these advancements in solving the problem of consciousness is still unclear. Based on technologies conceivably within the grasp of modern neuroscience, we discuss a thought experiment in which neural activity, in the form of action potentials, is initially recorded from all the neurons in a participant’s brain during a conscious experience and then played back into the same neurons. We consider whether this artificial replay can reconstitute a conscious experience. The possible outcomes of this experiment unravel hidden costs and pitfalls in understanding consciousness from the neurosciences’ perspective and challenge the conventional wisdom that causally links action potentials and consciousness.
Does brain activity cause consciousness? A thought experiment. https://doi.org/10.1371/journal.pbio.3001651.
Memory consolidation is a continuous transformative process between encoding and retrieval of mental representations. Recent research has shown that neural activity immediately after encoding is particularly associated with later successful retrieval. It is currently unclear whether post-encoding neural activity makes a distinct and causal contribution to memory consolidation. Here, we investigated the role of the post-encoding period for consolidation of spatial memory in neurologically normal human subjects. We used the GABAA-ergic anesthetic propofol to transiently manipulate neural activity during the initial stage of spatial memory consolidation without affecting encoding or retrieval. A total of 52 participants undergoing minor surgery learned to navigate to a target in a five-armed maze derived from animal experiments. Participants completed learning either immediately prior to injection of propofol (early group) or more than 60 min before injection (late group). Four hours after anesthesia, participants were tested for memory-guided navigation. Our results show a selective impairment of navigation in the early group and near-normal performance in the late group. Analysis of navigational error patterns further suggested that propofol impaired distinct aspects of spatial representations, in particular sequences of path segments and spatial relationships between landmarks. We conclude that neural activity during the post-encoding period makes a causal and specific contribution to consolidation of representations underlying self-centered and world-centered memory-guided navigation. Distinct aspects of these representations are susceptible to GABAA-ergic modulation within a post-encoding time-window of less than 60 min, presumably reflecting associative processes that contribute to the formation of integrated spatial representations that guide future behavior.
Post-encoding modulation of spatial memory consolidation by propofol. https://doi.org/10.1016/j.cortex.2022.08.004.
The use of head fixation has become routine in systems neuroscience. However, whether the behavior changes with head fixation, whether animals can learn aspects of a task while freely moving and transfer this knowledge to the head fixed condition, has not been examined in much detail. Here, we used a novel floating platform, the “Air-Track”, which simulates free movement in a real-world environment to address the effect of head fixation and developed methods to accelerate training of behavioral tasks for head fixed mice. We trained mice in a Y maze two choice discrimination task. One group was trained while head fixed and compared to a separate group that was pre-trained while freely moving and then trained on the same task while head fixed. Pre-training significantly reduced the time needed to relearn the discrimination task while head fixed. Freely moving and head fixed mice displayed similar behavioral patterns, however, head fixation significantly slowed movement speed. The speed of movement in the head fixed mice depended on the weight of the platform. We conclude that home-cage pre-training improves learning performance of head fixed mice and that while head fixation obviously limits some aspects of movement, the patterns of behavior observed in head fixed and freely moving mice are similar.
Efficient training approaches for optimizing behavioral performance and reducing head fixation time. https://doi.org/10.1371/journal.pone.0276531.
2021
Computer vision approaches have made significant inroads into offline tracking of behavior and estimating animal poses. In particular, because of their versatility, deep-learning approaches have been gaining attention in behavioral tracking without any markers. Here, we developed an approach using DeepLabCut for real-time estimation of movement. We trained a deep-neural network (DNN) offline with high-speed video data of a mouse whisking, then transferred the trained network to work with the same mouse, whisking in real-time. With this approach, we tracked the tips of three whiskers in an arc and converted positions into a TTL output within behavioral time scales, i.e., 10.5 ms. With this approach, it is possible to trigger output based on movement of individual whiskers, or on the distance between adjacent whiskers. Flexible closed-loop systems like the one we have deployed here can complement optogenetic approaches and can be used to directly manipulate the relationship between movement and neural activity.
Real-Time Closed-Loop Feedback in Behavioral Time Scales Using DeepLabCut. https://doi.org/10.1523/ENEURO.0415-20.2021.
Prompt execution of planned motor action is essential for survival. The interactions between frontal cortical circuits and the basal ganglia are central to goal-oriented action selection and initiation.1, 2, 3, 4 In rodents, the ventromedial thalamic nucleus (VM) is one of the critical nodes that conveys the output of the basal ganglia to the frontal cortical areas including the anterior lateral motor cortex (ALM).5, 6, 7, 8, 9 Recent studies showed the critical role of ALM and its interplay with the motor thalamus in preparing sensory-cued rewarded movements, specifically licking.10, 11, 12 Work in primates suggests that the basal ganglia output to the motor thalamus transmits an urgency or vigor signal,13, 14, 15 which leads to shortened reaction times and faster movement initiation. As yet, little is known about what signals are transmitted from the motor thalamus to the cortex during cued movements and how these signals contribute to movement initiation. In the present study, we employed a tactile-cued licking task in mice while monitoring reaction times of the initial lick. We found that inactivation of ALM delayed the initiation of cued licking. Two-photon Ca2+ imaging of VM axons revealed that the majority of the axon terminals in ALM were transiently active during licking. Their activity was predictive of the time of the first lick. Chemogenetic and optogenetic manipulation of VM axons in ALM indicated that VM inputs facilitate the initiation of cue-triggered and impulsive licking in trained mice. Our results suggest that VM thalamocortical inputs increase the probability and vigor of initiating planned motor responses.
Thalamic input to motor cortex facilitates goal-directed action initiation. https://doi.org/10.1016/j.cub.2021.06.089.
Synergistic interactions between independent synaptic input streams may fundamentally change the action potential (AP) output. Using partial information decomposition, we demonstrate here a substantial contribution of synergy between somatic and apical dendritic inputs to the information in the AP output of L5b pyramidal neurons. Activation of dendritic GABA B receptors (GABA B Rs), known to decrease APs in vivo , potently decreased synergy and increased somatic control of AP output. Synergy was the result of the voltage-dependence of the transfer resistance between dendrite and soma, which showed a two-fold increase per 28.7 mV dendritic depolarization. GIRK channels activated by dendritic GABA B Rs decreased voltage-dependent transfer resistances and AP output. In contrast, inhibition of dendritic L-type Ca 2+ channels prevented high-frequency bursts of APs, but did not affect dendro-somatic synergy. Finally, we show that NDNF-positive neurogliaform cells effectively control somatic AP via synaptic activation of dendritic GIRK channels. These results uncover a novel inhibitory mechanism that powerfully gates cellular information flow in the cortex.
GABAB Receptor-Mediated Regulation of Dendro-Somatic Synergy in Layer 5 Pyramidal Neurons. https://doi.org/10.3389/fncel.2021.718413.
Cortical layer 1 has a special role in long-term memory , Memories are generally considered to be embodied in the connection strengths between neurons. Finding the exact synaptic connections responsible for memories out of hundreds of trillions at first appears impossible. But new research may be taking us a step closer. Specific brain areas under the neocortex are known to be required for memory formation (including the hippocampus and basal ganglia), while long-term memory relating to facts and knowledge (semantic memory) is thought to reside in the neocortex ( 1 ). However, the precise location of semantic memory is still unknown, preventing researchers from understanding how and where memories are formed.
Memories off the top of your head. https://doi.org/10.1126/science.abk1859.
Intelligent behavior and cognitive functions in mammals depend on cortical microcircuits made up of a variety of excitatory and inhibitory cells that form a forest-like complex across six layers. Mechanistic understanding of cortical microcircuits requires both manipulation and monitoring of multiple layers and interactions between them. However, existing techniques are limited as to simultaneous monitoring and stimulation at different depths without damaging a large volume of cortical tissue. Here, we present a relatively simple and versatile method for delivering light to any two cortical layers simultaneously. The method uses a tiny optical probe consisting of two microprisms mounted on a single shaft. We demonstrate the versatility of the probe in three sets of experiments: first, two distinct cortical layers were optogenetically and independently manipulated; second, one layer was stimulated while the activity of another layer was monitored; third, the activity of thalamic axons distributed in two distinct cortical layers was simultaneously monitored in awake mice. Its simple-design, versatility, small-size, and low-cost allow the probe to be applied widely to address important biological questions.
Double-μPeriscope, a tool for multilayer optical recordings, optogenetic stimulations or both. https://doi.org/10.7554/eLife.72894.
Optical imaging techniques are widely used in biological research, but their penetration depth is limited by tissue scattering. Wavefront shaping techniques are able to overcome this problem in principle, but are often slow, and their performance depends on the sample. This greatly reduces their practicability for biological applications. Here we present a scattering compensation technique based on three-photon excitation, which converges faster than comparable two-photon (2P) techniques and works reliably even on densely labeled samples, where 2P approaches fail. To demonstrate its usability and advantages for biomedical imaging, we apply it to the imaging of dendritic spines on layer 5 neurons labeled with green fluorescent protein in an anesthetized mouse.
Deep tissue scattering compensation with three-photon F-SHARP. https://doi.org/10.1364/OPTICA.440279.
2020
Human dendrites are special A special developmental program in the human brain drives the disproportionate thickening of cortical layer 2/3. This suggests that the expansion of layer 2/3, along with its numerous neurons and their large dendrites, may contribute to what makes us human. Gidon et al. thus investigated the dendritic physiology of layer 2/3 pyramidal neurons in slices taken from surgically resected brain tissue in epilepsy patients. Dual somatodendritic recordings revealed previously unknown classes of action potentials in the dendrites of these neurons, which make their activity far more complex than has been previously thought. These action potentials allow single neurons to solve two long-standing computational problems in neuroscience that were considered to require multilayer neural networks. Science , this issue p. 83 , Dendritic action potentials extend the repertoire of computations available to human neurons. , The active electrical properties of dendrites shape neuronal input and output and are fundamental to brain function. However, our knowledge of active dendrites has been almost entirely acquired from studies of rodents. In this work, we investigated the dendrites of layer 2 and 3 (L2/3) pyramidal neurons of the human cerebral cortex ex vivo. In these neurons, we discovered a class of calcium-mediated dendritic action potentials (dCaAPs) whose waveform and effects on neuronal output have not been previously described. In contrast to typical all-or-none action potentials, dCaAPs were graded; their amplitudes were maximal for threshold-level stimuli but dampened for stronger stimuli. These dCaAPs enabled the dendrites of individual human neocortical pyramidal neurons to classify linearly nonseparable inputs—a computation conventionally thought to require multilayered networks.
Dendritic action potentials and computation in human layer 2/3 cortical neurons. https://doi.org/10.1126/science.aax6239.
The mystery of general anesthesia is that it specifically suppresses consciousness by disrupting feedback signaling in the brain, even when feedforward signaling and basic neuronal function are left relatively unchanged. The mechanism for such selectiveness is unknown. Here we show that three different anesthetics have the same disruptive influence on signaling along apical dendrites in cortical layer 5 pyramidal neurons in mice. We found that optogenetic depolarization of the distal apical dendrites caused robust spiking at the cell body under awake conditions that was blocked by anesthesia. Moreover, we found that blocking metabotropic glutamate and cholinergic receptors had the same effect on apical dendrite decoupling as anesthesia or inactivation of the higher-order thalamus. If feedback signaling occurs predominantly through apical dendrites, the cellular mechanism we found would explain not only how anesthesia selectively blocks this signaling but also why conscious perception depends on both cortico-cortical and thalamo-cortical connectivity.
General Anesthesia Decouples Cortical Pyramidal Neurons. https://doi.org/10.1016/j.cell.2020.01.024.
Layer 6b (L6b), the deepest neocortical layer, projects to cortical targets and higher-order thalamus and is the only layer responsive to the wake-promoting neuropeptide orexin/hypocretin. These characteristics suggest that L6b can strongly modulate brain state, but projections to L6b and their influence remain unknown. Here, we examine the inputs to L6b ex vivo in the mouse primary somatosensory cortex with rabies-based retrograde tracing and channelrhodopsin-assisted circuit mapping in brain slices. We find that L6b receives its strongest excitatory input from intracortical long-range projection neurons, including those in the contralateral hemisphere. In contrast, local intracortical input and thalamocortical input were significantly weaker. Moreover, our data suggest that L6b receives far less thalamocortical input than other cortical layers. L6b was most strongly inhibited by PV and SST interneurons. This study shows that L6b integrates long-range intracortical information and is not part of the traditional thalamocortical loop.
Layer 6b Is Driven by Intracortical Long-Range Projection Neurons. https://doi.org/10.1016/j.celrep.2020.02.044.
In the course of a day, brain states fluctuate, from conscious awake information-acquiring states to sleep states, during which previously acquired information is further processed and stored as memories. One hypothesis is that memories are consolidated and stored during “offline” states such as sleep, a process thought to involve transfer of information from the hippocampus to other cortical areas. Up and Down states (UDS), patterns of activity that occur under anesthesia and sleep states, are likely to play a role in this process, although the nature of this role remains unclear. Here we review what is currently known about these mechanisms in three anatomically distinct but interconnected cortical areas: somatosensory cortex, entorhinal cortex, and the hippocampus. In doing so, we consider the role of this activity in the coordination of “replay” during sleep states, particularly during hippocampal sharp-wave ripples. We conclude that understanding the generation and propagation of UDS may provide key insights into the cortico-hippocampal dialogue linking archi- and neocortical areas during memory formation.
Up and Down States and Memory Consolidation Across Somatosensory, Entorhinal, and Hippocampal Cortices. https://doi.org/10.3389/fnsys.2020.00022.
In this review article, we highlight several disparate ideas that are linked to changes in brain state (i.e., sleep to arousal, Down to Up, synchronized to de-synchronized). In any discussion of the brain state, we propose that the cortical pyramidal neuron has a central position. EEG recordings, which typically assess brain state, predominantly reflect the activity of cortical pyramidal neurons. This means that the dominant rhythmic activity that characterizes a particular brain state ultimately has to manifest globally across the pyramidal neuron population. During state transitions, it is the long-range connectivity of these neurons that broadcast the resultant changes in activity to many subcortical targets. Structures like the thalamus, brainstem/hypothalamic neuromodulatory systems, and respiratory systems can also strongly influence brain state, and for many decades we have been uncovering bidirectional pathways that link these structures to state changes in the cerebral cortex. More recently, movement and active behaviors have emerged as powerful drivers of state changes. Each of these systems involve different circuits distributed across the brain. Yet, for a system-wide change in brain state, there must be a collaboration between these circuits that reflects and perhaps triggers the transition between brain states. As we expand our understanding of how brain state changes, our current challenge is to understand how these diverse sets of circuits and pathways interact to produce the changes observed in cortical pyramidal neurons.
Perspective on the Multiple Pathways to Changing Brain States. https://doi.org/10.3389/fnsys.2020.00023.
Abstract Optical microscopy is an indispensable tool in biomedical sciences, but its reach in deep tissues is limited due to aberrations and scattering. This problem can be overcome by wavefront-shaping techniques, albeit at limited fields of view (FOVs). Inspired by astronomical imaging, conjugate wavefront shaping can lead to an increased field of view in microscopy, but this correction is limited to a set depth and cannot be dynamically adapted. Here, we present a conjugate wavefront-shaping scheme based on focus scanning holographic aberration probing (F-SHARP). We combine it with a compact implementation that can be readily adapted to a variety of commercial and home-built two-photon microscopes. We demonstrate the power of the method by imaging with high resolution over extended FOV (>80 µm) deeper than 400 μm inside a mouse brain through a thinned skull.
Dynamic conjugate F-SHARP microscopy. https://doi.org/10.1038/s41377-020-00348-x.
Active dendritic currents gate descending cortical outputs in perception. https://doi.org/10.1038/s41593-020-0677-8.
Recent breakthroughs in neurobiology indicate that the time is ripe to understand how cellular-level mechanisms are related to conscious experience. Here, we highlight the biophysical properties of pyramidal cells, which allow them to act as gates that control the evolution of global activation patterns. In conscious states, this cellular mechanism enables complex sustained dynamics within the thalamocortical system, whereas during unconscious states, such signal propagation is prohibited. We suggest that the hallmark of conscious processing is the flexible integration of bottom-up and top-down data streams at the cellular level. This cellular integration mechanism provides the foundation for Dendritic Integration Theory, a novel neurobiological theory of consciousness.
Cellular Mechanisms of Conscious Processing. https://doi.org/10.1016/j.tics.2020.07.006.
Memory consolidation in the neocortex Information transfer between brain structures located in the medial-temporal lobe and the neocortex is essential for learning. However, the neuronal underpinnings of this transfer are unknown. Doron et al. found that neurons located in the deep layers of the perirhinal cortex exhibit increased firing after microstimulation upon learning (see the Perspective by Donato). Learning was associated with the emergence of a small population of neurons in layer 5 of the somatosensory cortex that increased bursting upon stimulation. This increase in bursting was accompanied by an increase in dendritic activity, and silencing the perirhinal cortex to layer 1 projection effectively disrupted learning and its physiological correlates. During learning, perirhinal inputs thus act as a gate for the enhancement of cortico-cortical inputs, which are necessary for stimulus detection and are strengthened during learning. Science , this issue p. eaaz3136 ; see also p. 1410 , Perirhinal input, predominantly to sensory cortical layer 1, controls hippocampal-dependent associative learning in mice. , INTRODUCTION Arguably one of the biggest mysteries in neuroscience is how the brain stores long-term memories. Since the 1950s, it has been well established that long-term memories reside in the neocortex but that their formation is dependent on the hippocampus and medial-temporal lobe structures. It is therefore remarkable that we still do not know what cellular mechanisms underlie long-term memory storage in the neocortex or exactly where they are located. RATIONALE The primary challenge for investigating the neural circuit underlying memory formation in the neocortex is the distributed nature of the resulting memory trace throughout the cortex. We therefore used a behavioral paradigm dependent on both the hippocampus and neocortex that enabled us to generate memory traces in a specific cortical location by training rodents to associate the direct electrical microstimulation of the cortex with a reward. This also allowed us to specifically examine the contribution of circuit elements in the defined anatomical location. Rodents learned to behaviorally report the microstimulation within only a few trials and improved over a 3-day period, during which we examined the evolution of learned neuronal responses in the stimulated area. We hypothesized that the influence of the hippocampus on memory formation in the neocortex occurs at the interface between these two structures. RESULTS We first confirmed that learning to associate microstimulation of the primary somatosensory cortex (S1) with a reward depends on hippocampal activity using suppression of action potentials (APs) with lidocaine in this brain area. Using retrograde and anterograde tracing methods, we found that the perirhinal cortex (the last station in the medial-temporal loop projecting to S1) predominantly targets layer 1 (L1), suggesting that important events relating to memory formation occur in neocortical L1. We tested this with targeted chemogenetic suppression of perirhinal input to L1 above the stimulated S1 region. Notably, this very specific and localized manipulation was sufficient to disrupt learning. The effect was learning-specific and had no influence in expert animals, which demonstrated that the perirhinal input did not alter the ability to perceive and behaviorally report the stimulus, per se. We found that the perirhinal cortex signaled information related to successful behavior during learning, gated the evolution of distinct firing, and enhanced burst responses in 11% of layer 5 (L5) pyramidal neurons in S1 (40% of neurons had reduced firing responses and 49% showed no change in firing). Apical dendritic excitability was correspondingly enhanced in a similar proportion of L5 pyramidal neurons. This suggested that the mechanism for memory formation had a dendritic origin. Consistent with this hypothesis, we found that both activation of γ-aminobutyric acid type B (GABA B ) receptors—which disrupt apical dendritic calcium (Ca 2+ ) activity—and activation of dendritic-targeting, somatostatin-positive interneurons impaired memory formation similarly to suppressing perirhinal input to L1. Finally, we found that after learning the microstimulation detection task, evoking a single burst of APs (but not the same number of low-frequency spikes) in a single L5 pyramidal neuron could trigger behavior. CONCLUSION We found that medial-temporal input to neocortical L1 gates the evolution of specific firing responses in subpopulations of L5 pyramidal neurons including up- and down-regulated firing patterns and an elevation in burstiness by means of a mechanism that is most likely related to apical dendritic activity. After learning, these neocortical responses become independent of the medial-temporal influence but continue to evoke behavior with bursts conveying higher saliency. We conclude that L1 is the locus for hippocampal-dependent associative learning in the neocortex, where memory engrams are established in subsets of pyramidal neurons by enhancing the sensitivity of tuft dendrites to contextual inputs and driving burst firing. Probing the influence of the medial-temporal lobe on memory formation. ( A ) The perirhinal cortex (PRh), part of the medial-temporal lobe (MTL) structures (blue box), targets L1 in S1. ( B ) We performed microstimulation (μStim) of L5 in the sensory cortex while chemogenetically suppressing the axonal projection to cortical L1 (red) from PRh (blue arrow). ( C ) Rodents learned to associate μStim with water reward. ( D to F ) Learning was suppressed by blocking MTL input [(D) and (E)] that otherwise evoked dendritic activity in L5 pyramidal neurons [(F), orange tuft], which allowed them to associate contextual input to L1 with the μStim. ( G ) More than 3 days of learning led to subpopulations of firing responses in L5 cells in S1 such that evoking a burst but not low-frequency spikes in single L5 cells retrieved learned behavior. , Hippocampal output influences memory formation in the neocortex, but this process is poorly understood because the precise anatomical location and the underlying cellular mechanisms remain elusive. Here, we show that perirhinal input, predominantly to sensory cortical layer 1 (L1), controls hippocampal-dependent associative learning in rodents. This process was marked by the emergence of distinct firing responses in defined subpopulations of layer 5 (L5) pyramidal neurons whose tuft dendrites receive perirhinal inputs in L1. Learning correlated with burst firing and the enhancement of dendritic excitability, and it was suppressed by disruption of dendritic activity. Furthermore, bursts, but not regular spike trains, were sufficient to retrieve learned behavior. We conclude that hippocampal information arriving at L5 tuft dendrites in neocortical L1 mediates memory formation in the neocortex.
Perirhinal input to neocortical layer 1 controls learning. https://doi.org/10.1126/science.aaz3136.
2019
A central function of the brain is to plan, predict, and imagine the effect of movement in a dynamically changing environment. Here we show that in mice head-fixed in a plus-maze, floating on air, and trained to pick lanes based on visual stimuli, the asymmetric movement, and position of whiskers on the two sides of the face signals whether the animal is moving, turning, expecting reward, or licking. We show that (1) whisking asymmetry is coordinated with behavioral state, and that behavioral state can be decoded and predicted based on asymmetry, (2) even in the absence of tactile input, whisker positioning and asymmetry nevertheless relate to behavioral state, and (3) movement of the nose correlates with asymmetry, indicating that facial expression of the mouse is itself correlated with behavioral state. These results indicate that the movement of whiskers, a behavior that is not instructed or necessary in the task, can inform an observer about what a mouse is doing in the maze. Thus, the position of these mobile tactile sensors reflects a behavioral and movement-preparation state of the mouse. SIGNIFICANCE STATEMENT Behavior is a sequence of movements, where each movement can be related to or can trigger a set of other actions. Here we show that, in mice, the movement of whiskers (tactile sensors used to extract information about texture and location of objects) is coordinated with and predicts the behavioral state of mice: that is, what mice are doing, where they are in space, and where they are in the sequence of behaviors.
Whisking Asymmetry Signals Motor Preparation and the Behavioral State of Mice. https://doi.org/10.1523/JNEUROSCI.1809-19.2019.
The advent of optogenetic methods has made it possible to use endogeneously produced molecules to image and manipulate cellular, subcellular, and synaptic activity. It has also led to the development of photoactivatable calcium-dependent indicators that mark active synapses, neurons, and circuits. Furthermore, calcium-dependent photoactivation can be used to trigger gene expression in active neurons. Here we describe two sets of protocols, one using CaMPARI and a second one using Cal-Light. CaMPARI, a calcium-modulated photoactivatable ratiometric integrator, enables rapid network-wide, tunable, all-optical functional circuit mapping. Cal-Light, a photoactivatable calcium sensor, while slower to respond than CaMPARI, has the capacity to trigger the expression of genes, including effectors, activators, indicators, or other constructs. Here we describe the rationale and provide procedures for using these two calcium-dependent constructs (1) in vitro in dissociated primary neuronal cell cultures (CaMPARI & Cal-Light); (2) in vitro in acute brain slices for circuit mapping (CaMPARI); (3) in vivo for triggering photoconversion or gene expression (CaMPARI & Cal-Light); and finally, (4) for recovering photoconverted neurons post-fixation with immunocytochemistry (CaMPARI). The approaches and protocols we describe are examples of the potential uses of both CaMPARI & Cal-Light. The ability to mark and manipulate neurons that are active during specific epochs of behavior has a vast unexplored experimental potential.
Optically Induced Calcium-Dependent Gene Activation and Labeling of Active Neurons Using CaMPARI and Cal-Light. https://doi.org/10.3389/fnsyn.2019.00016.
Oxytocin (OT) release by axonal terminals onto the central nucleus of the amygdala exerts anxiolysis. To investigate which subpopulation of OT neurons contributes to this effect, we developed a novel method: virus-delivered genetic activity-induced tagging of cell ensembles (vGATE). With the vGATE method, we identified and permanently tagged a small subpopulation of OT cells, which, by optogenetic stimulation, strongly attenuated contextual fear-induced freezing, and pharmacogenetic silencing of tagged OT neurons impaired context-specific fear extinction, demonstrating that the tagged OT neurons are sufficient and necessary, respectively, to control contextual fear. Intriguingly, OT cell terminals of fear-experienced rats displayed enhanced glutamate release in the amygdala. Furthermore, rats exposed to another round of fear conditioning displayed 5-fold more activated magnocellular OT neurons in a novel environment than a familiar one, possibly for a generalized fear response. Thus, our results provide first evidence that hypothalamic OT neurons represent a fear memory engram.
A Fear Memory Engram and Its Plasticity in the Hypothalamic Oxytocin System. https://doi.org/10.1016/j.neuron.2019.04.029.
The topographic map in layer 4 of somatosensory cortex is usually specified early postnatally and stable thereafter. Genital cortex, however, undergoes a sex-hormone- and sexual-touch-dependent pubertal expansion. Here, we image pubertal development of genital cortex in Scnn1a-Tg3-Cre mice, where transgene expression has been shown to be restricted to layer 4 neurons with primary sensory cortex identity. Interestingly, during puberty, the number of Scnn1a+ neurons roughly doubled within genital cortex. The increase of Scnn1a+ neurons was gradual and rapidly advanced by initial sexual experience. Neurons that gained Scnn1a expression comprised stellate and pyramidal neurons in layer 4. Unlike during neonatal development, pyramids did not retract their apical dendrites during puberty. Calcium imaging revealed stronger genital-touch responses in Scnn1a+ neurons in males versus females and a developmental increase in responsiveness in females. The first sexual interaction is a unique physical experience that often creates long-lasting memories. We suggest such experience uniquely alters somatosensory body maps.
Effects of Sexual Experience and Puberty on Mouse Genital Cortex revealed by Chronic Imaging. https://doi.org/10.1016/j.cub.2019.08.062.
One fundamental feature of consciousness is that the contents of consciousness depend on the state of consciousness. Here, we propose an answer to why this is so: both the state and the contents of consciousness depend on the activity of cortical layer 5 pyramidal (L5p) neurons. These neurons affect both cortical and thalamic processing, hence coupling the cortico-cortical and thalamo-cortical loops with each other. Functionally this coupling corresponds to the coupling between the state and the contents of consciousness. Together the cortico-cortical and thalamo-cortical loops form a thalamo-cortical broadcasting system, where the L5p cells are the central elements. This perspective makes one quite specific prediction: cortical processing that does not include L5p neurons will be unconscious. More generally, the present perspective suggests that L5p neurons have a central role in the mechanisms underlying consciousness.
Coupling the State and Contents of Consciousness. https://doi.org/10.3389/fnsys.2019.00043.
2018
This review article addresses the function of the layers of the cerebral cortex. We develop the perspective that cortical layering needs to be understood in terms of its functional anatomy, i.e., the terminations of synaptic inputs on distinct cellular compartments and their effect on cortical activity. The cortex is a hierarchical structure in which feed forward and feedback pathways have a layer-specific termination pattern. We take the view that the influence of synaptic inputs arriving at different cortical layers can only be understood in terms of their complex interaction with cellular biophysics and the subsequent computation that occurs at the cellular level. We use high-resolution fMRI, which can resolve activity across layers, as a case study for implementing this approach by describing how cognitive events arising from the laminar distribution of inputs can be interpreted by taking into account the properties of neurons that span different layers. This perspective is based on recent advances in measuring subcellular activity in distinct feed-forward and feedback axons and in dendrites as they span across layers.
A Perspective on Cortical Layering and Layer-Spanning Neuronal Elements. https://doi.org/10.3389/fnana.2018.00056.
Abstract Marking functionally distinct neuronal ensembles with high spatiotemporal resolution is a key challenge in systems neuroscience. We recently introduced CaMPARI, an engineered fluorescent protein whose green-to-red photoconversion depends on simultaneous light exposure and elevated calcium, which enabled marking active neuronal populations with single-cell and subsecond resolution. However, CaMPARI (CaMPARI1) has several drawbacks, including background photoconversion in low calcium, slow kinetics and reduced fluorescence after chemical fixation. In this work, we develop CaMPARI2, an improved sensor with brighter green and red fluorescence, faster calcium unbinding kinetics and decreased photoconversion in low calcium conditions. We demonstrate the improved performance of CaMPARI2 in mammalian neurons and in vivo in larval zebrafish brain and mouse visual cortex. Additionally, we herein develop an immunohistochemical detection method for specific labeling of the photoconverted red form of CaMPARI. The anti-CaMPARI-red antibody provides strong labeling that is selective for photoconverted CaMPARI in activated neurons in rodent brain tissue.
Improved methods for marking active neuron populations. https://doi.org/10.1038/s41467-018-06935-2.
2017
Abstract Here, we describe an automated optical method for tracking animal behavior in both head-fixed and freely moving animals, in real time and offline. It takes advantage of an off-the-shelf camera system, the Pixy camera, designed as a fast vision sensor for robotics that uses a color-based filtering algorithm at 50 Hz to track objects. Using customized software, we demonstrate the versatility of our approach by first tracking the rostro-caudal motion of individual adjacent row (D1, D2) or arc whiskers (β, γ), or a single whisker and points on the whisker pad, in head-fixed mice performing a tactile task. Next, we acquired high-speed video and Pixy data simultaneously and applied the pixy-based real-time tracking to high-speed video data. With this approach, we expand the temporal resolution of the Pixy camera and track motion ( post hoc ) at the limit of high-speed video frame rates. Finally, we show that this system is flexible: it can be used to track individual whisker or limb position without any sophisticated object tracking algorithm, it can be used in many lighting conditions including infrared (IR); it can be used to track head rotation and location of multiple animals simultaneously. Our system makes behavioral monitoring possible in virtually any biological setting.
Pixying Behavior: A Versatile Real-Time and <i>Post Hoc</i> Automated Optical Tracking Method for Freely Moving and Head Fixed Animals. https://doi.org/10.1523/ENEURO.0245-16.2017.
A Reward-Based Behavioral Platform to Measure Neural Activity during Head-Fixed Behavior. https://doi.org/10.3389/fncel.2017.00156.
Key points The genetically encoded fluorescent calcium integrator calcium‐modulated photoactivatable ratiobetric integrator (CaMPARI) reports calcium influx induced by synaptic and neural activity. Its fluorescence is converted from green to red in the presence of violet light and calcium. The rate of conversion – the sensitivity to activity – is tunable and depends on the intensity of violet light. Synaptic activity and action potentials can independently initiate significant CaMPARI conversion. The level of conversion by subthreshold synaptic inputs is correlated to the strength of input, enabling optical readout of relative synaptic strength. When combined with optogenetic activation of defined presynaptic neurons, CaMPARI provides an all‐optical method to map synaptic connectivity. Abstract The calcium‐modulated photoactivatable ratiometric integrator (CaMPARI) is a genetically encoded calcium integrator that facilitates the study of neural circuits by permanently marking cells active during user‐specified temporal windows. Permanent marking enables measurement of signals from large swathes of tissue and easy correlation of activity with other structural or functional labels. One potential application of CaMPARI is labelling neurons postsynaptic to specific populations targeted for optogenetic stimulation, giving rise to all‐optical functional connectivity mapping. Here, we characterized the response of CaMPARI to several common types of neuronal calcium signals in mouse acute cortical brain slices. Our experiments show that CaMPARI is effectively converted by both action potentials and subthreshold synaptic inputs, and that conversion level is correlated to synaptic strength. Importantly, we found that conversion rate can be tuned: it is linearly related to light intensity. At low photoconversion light levels CaMPARI offers a wide dynamic range due to slower conversion rate; at high light levels conversion is more rapid and more sensitive to activity. Finally, we employed CaMPARI and optogenetics for functional circuit mapping in ex vivo acute brain slices, which preserve in vivo ‐like connectivity of axon terminals. With a single light source, we stimulated channelrhodopsin‐2‐expressing long‐range posteromedial (POm) thalamic axon terminals in cortex and induced CaMPARI conversion in recipient cortical neurons. We found that POm stimulation triggers robust photoconversion of layer 5 cortical neurons and weaker conversion of layer 2/3 neurons. Thus, CaMPARI enables network‐wide, tunable, all‐optical functional circuit mapping that captures supra‐ and subthreshold depolarization. , Key points The genetically encoded fluorescent calcium integrator calcium‐modulated photoactivatable ratiobetric integrator (CaMPARI) reports calcium influx induced by synaptic and neural activity. Its fluorescence is converted from green to red in the presence of violet light and calcium. The rate of conversion – the sensitivity to activity – is tunable and depends on the intensity of violet light. Synaptic activity and action potentials can independently initiate significant CaMPARI conversion. The level of conversion by subthreshold synaptic inputs is correlated to the strength of input, enabling optical readout of relative synaptic strength. When combined with optogenetic activation of defined presynaptic neurons, CaMPARI provides an all‐optical method to map synaptic connectivity.
All‐optical functional synaptic connectivity mapping in acute brain slices using the calcium integrator CaMPARI. https://doi.org/10.1113/JP273116.
Abstract Cortical surface recording techniques such as EEG and ECoG are widely used for measuring brain activity. The prevailing assumption is that surface potentials primarily reflect synaptic activity, although non-synaptic events may also contribute. Here we show that dendritic calcium spikes occurring in pyramidal neurons (that we showed previously are cognitively relevant) are clearly detectable in cortical surface potentials. To show this we developed an optogenetic, non-synaptic approach to evoke dendritic calcium spikes in vivo. We found that optogenetically evoked calcium spikes were easily detectable and had an unexpected waveform near the cortical surface. Sensory-evoked dendritic calcium spikes were also clearly detectable with amplitudes that matched the contribution of synaptic input. These results reveal how dendritic calcium spikes appear at the cortical surface and their significant impact on surface potentials, suggesting that long-standing surface recording data may contain information about dendritic activity that is relevant to behavior and cognitive function.
Dendritic calcium spikes are clearly detectable at the cortical surface. https://doi.org/10.1038/s41467-017-00282-4.
Abstract How sleep influences brain plasticity is not known. In particular, why certain electroencephalographic (EEG) rhythms are linked to memory consolidation is poorly understood. Calcium activity in dendrites is known to be necessary for structural plasticity changes, but this has never been carefully examined during sleep. Here, we report that calcium activity in populations of neocortical dendrites is increased and synchronised during oscillations in the spindle range in naturally sleeping rodents. Remarkably, the same relationship is not found in cell bodies of the same neurons and throughout the cortical column. Spindles during sleep have been suggested to be important for brain development and plasticity. Our results provide evidence for a physiological link of spindles in the cortex specific to dendrites, the main site of synaptic plasticity.
Cortical dendritic activity correlates with spindle-rich oscillations during sleep in rodents. https://doi.org/10.1038/s41467-017-00735-w.
What can artificial intelligence learn from neuroscience, and vice versa?
Branching into brains. https://doi.org/10.7554/eLife.33066.
Science is ideally suited to connect people from different cultures and thereby foster mutual understanding. To promote international life science collaboration, we have launched “The Science Bridge” initiative. Our current project focuses on partnership between Western and Middle Eastern neuroscience communities.
Building Bridges through Science. https://doi.org/10.1016/j.neuron.2017.09.028.
2016
Natural behavior occurs in multiple sensory and motor modalities and in particular is dependent on sensory feedback that constantly adjusts behavior. To investigate the underlying neuronal correlates of natural behavior, it is useful to have access to state-of-the-art recording equipment (e.g., 2-photon imaging, patch recordings, etc.) that frequently requires head fixation. This limitation has been addressed with various approaches such as virtual reality/air ball or treadmill systems. However, achieving multimodal realistic behavior in these systems can be challenging. These systems are often also complex and expensive to implement. Here we present “Air-Track,” an easy-to-build head-fixed behavioral environment that requires only minimal computational processing. The Air-Track is a lightweight physical maze floating on an air table that has all the properties of the “real” world, including multiple sensory modalities tightly coupled to motor actions. To test this system, we trained mice in Go/No-Go and two-alternative forced choice tasks in a plus maze. Mice chose lanes and discriminated apertures or textures by moving the Air-Track back and forth and rotating it around themselves. Mice rapidly adapted to moving the track and used visual, auditory, and tactile cues to guide them in performing the tasks. A custom-controlled camera system monitored animal location and generated data that could be used to calculate reaction times in the visual and somatosensory discrimination tasks. We conclude that the Air-Track system is ideal for eliciting natural behavior in concert with virtually any system for monitoring or manipulating brain activity.
Air-Track: a real-world floating environment for active sensing in head-fixed mice. https://doi.org/10.1152/jn.00088.2016.
One of the leading approaches to non-invasively treat a variety of brain disorders is transcranial magnetic stimulation (TMS). However, despite its clinical prevalence, very little is known about the action of TMS at the cellular level let alone what effect it might have at the subcellular level (e.g. dendrites). Here, we examine the effect of single-pulse TMS on dendritic activity in layer 5 pyramidal neurons of the somatosensory cortex using an optical fiber imaging approach. We find that TMS causes GABAB-mediated inhibition of sensory-evoked dendritic Ca2+ activity. We conclude that TMS directly activates fibers within the upper cortical layers that leads to the activation of dendrite-targeting inhibitory neurons which in turn suppress dendritic Ca2+ activity. This result implies a specificity of TMS at the dendritic level that could in principle be exploited for investigating these structures non-invasively. , The brain’s billions of neurons communicate with one another using electrical signals. Applying a magnetic field to a small area of the scalp can temporarily disrupt these signals by inducing small electrical currents in the brain tissue underneath. The currents interfere with the brain’s own electrical signals and temporarily disrupt the activity of the stimulated brain region. This technique, which is known as transcranial magnetic stimulation, is often used to investigate the roles of specific brain regions. By examining what happens when a region is briefly taken ‘offline’, it is possible to deduce what that area normally does. Transcranial magnetic stimulation is also used to treat brain disorders ranging from epilepsy to schizophrenia without the need for surgery or drugs. But despite its widespread usage, little is known about how transcranial magnetic stimulation affects individual neurons. Neurons are made up of a cell body, which has numerous short branches called dendrites, and a cable-like structure called the axon. Neurons signal to each other by releasing chemical messengers across junctions called synapses. The chemical signals are generally released from the axon of one neuron and bind to receptor proteins on a dendrite on another neuron to stimulate electrical activity in the receiving neuron. Murphy et al. have now investigated how transcranial magnetic stimulation affects the activity of dendrites from neurons within the cortex of the rat brain. This revealed that the magnetic fields stimulate other neurons that inhibit the activity of dendrites from neurons within the deeper layers of the cortex. The inhibition process depends on a type of receptor protein in the dendrites called GABAB receptors; blocking these receptors prevents transcranial magnetic stimulation from altering the activity of stimulated brain regions. The processes occurring in these dendrites have been linked to cognitive function. The next challenge will be to integrate the non-invasive transcranial magnetic stimulation approach with cognitive tests in humans that can now manipulate dendritic activity to test their importance under various circumstances.
Transcranial magnetic stimulation (TMS) inhibits cortical dendrites. https://doi.org/10.7554/eLife.13598.
Neocortical pyramidal cells can integrate two classes of input separately and use one to modulate response to the other. Their tuft dendrites are electrotonically separated from basal dendrites and soma by the apical dendrite, and apical hyperpolarization-activated currents (Ih) further isolate subthreshold integration of tuft inputs. When apical depolarization exceeds a threshold, however, it can enhance response to the basal inputs that specify the cell’s selective sensitivity. This process is referred to as apical amplification (AA). We review evidence suggesting that, by regulating Ih in the apical compartments, adrenergic arousal controls the coupling between apical and somatic integration zones thus modifying cognitive capabilities closely associated with consciousness. Evidence relating AA to schizophrenia, sleep, and anesthesia is reviewed, and we assess theories that emphasize the relevance of AA to consciousness. Implications for theories of neocortical computation that emphasize context-sensitive modulation are summarized. We conclude that the findings concerning AA and its regulation by arousal offer a new perspective on states of consciousness, the function and evolution of neocortex, and psychopathology. Many issues worthy of closer examination arise.
The effects of arousal on apical amplification and conscious state. https://doi.org/10.1093/nc/niw015.
Now you feel it, now you don't What determines the detection of a sensory stimulus? To address this question, Takahashi et al. combined in vivo two-photon imaging, electrophysiology, optogenetics, and behavioral analysis in a study of mice. Calcium signals in apical dendrites of pyramidal neurons in the somatosensory cortex controlled the perceptual threshold of the mice's whiskers. Strong reduction of dendritic calcium signaling impaired the perceptual detection threshold so that an identical stimulus could no longer be noticed. Abstract There is as yet no consensus concerning the neural basis of perception and how it operates at a mechanistic level. We found that Ca2+ activity in the apical dendrites of a subset of layer 5 (L5) pyramidal neurons in primary somatosensory cortex (S1) in mice is correlated with the threshold for perceptual detection of whisker deflections. Manipulating the activity of apical dendrites shifted the perceptual threshold, demonstrating that an active dendritic mechanism is causally linked to perceptual detection.
Active cortical dendrites modulate perception. https://doi.org/10.1126/science.aah6066.
2015
A fundamental issue in cortical processing of sensory information is whether top-down control circuits from higher brain areas to primary sensory areas not only modulate but actively engage in perception. Here, we report the identification of a neural circuit for top-down control in the mouse somatosensory system. The circuit consisted of a long-range reciprocal projection between M2 secondary motor cortex and S1 primary somatosensory cortex. In vivo physiological recordings revealed that sensory stimulation induced sequential S1 to M2 followed by M2 to S1 neural activity. The top-down projection from M2 to S1 initiated dendritic spikes and persistent firing of S1 layer 5 (L5) neurons. Optogenetic inhibition of M2 input to S1 decreased L5 firing and the accurate perception of tactile surfaces. These findings demonstrate that recurrent input to sensory areas is essential for accurate perception and provide a physiological model for one type of top-down control circuit.
A Top-Down Cortical Circuit for Accurate Sensory Perception. https://doi.org/10.1016/j.neuron.2015.05.006.
L5 pyramidal neurons are the only neocortical cell type with dendrites reaching all six layers of cortex, casting them as one of the main integrators in the cortical column. What is the nature and mode of computation performed in mouse primary visual cortex (V1) given the physiology of L5 pyramidal neurons? First, we experimentally establish active properties of the dendrites of L5 pyramidal neurons of mouse V1 using patch-clamp recordings. Using a detailed multi-compartmental model, we show this physiological setup to be well suited for coincidence detection between basal and apical tuft inputs by controlling the frequency of spike output. We further show how direct inhibition of calcium channels in the dendrites modulates such coincidence detection. To establish the singe-cell computation that this biophysics supports, we show that the combination of frequency-modulation of somatic output by tuft input and (simulated) calcium-channel blockage functionally acts as a composite sigmoidal function. Finally, we explore how this computation provides a mechanism whereby dendritic spiking contributes to orientation tuning in pyramidal neurons.
Physiology of Layer 5 Pyramidal Neurons in Mouse Primary Visual Cortex: Coincidence Detection through Bursting. https://doi.org/10.1371/journal.pcbi.1004090.
Design and performance of an ultra-flexible two-photon microscope for in vivo research. https://doi.org/10.1364/BOE.6.004228.
2014
Recent evidence in vitro suggests that the tuft dendrites of pyramidal neurons are capable of evoking local NMDA receptor-dependent electrogenesis, so-called NMDA spikes. However, it has so far proved difficult to demonstrate their existence in vivo. Moreover, it is not clear whether NMDA spikes are relevant to the output of pyramidal neurons. We found that local NMDA spikes occurred in tuft dendrites of layer 2/3 pyramidal neurons both spontaneously and following sensory input, and had a large influence on the number of output action potentials. Using two-photon activation of an intracellular caged NMDA receptor antagonist (tc-MK801), we found that isolated NMDA spikes typically occurred in multiple branches simultaneously and that sensory stimulation substantially increased their probability. Our results demonstrate that NMDA receptors have a vital role in coupling the tuft region of the layer 2/3 pyramidal neuron to the cell body, enhancing the effectiveness of layer 1 input.
NMDA spikes enhance action potential generation during sensory input. https://doi.org/10.1038/nn.3646.
Translating the advances seen recently in recording from neurons in vivo to dendritic recordings presents special difficulties. In vivo two-photon imaging of dendrites was achieved over a decade ago and is still the method of choice for recording from small dendritic compartments in single neurons but has proven more difficult to apply to many dendrites simultaneously or to awake, freely moving preparations. An alternative that can be applied to layer 5 neocortical pyramidal neurons is the use of a fiber-optic method combined with bolus loading of fluorescent calcium indicator. This method takes advantage of the fact that the apical dendrites of these neurons are long (~1 mm) and always project in the same manner towards the cortical surface, thus allowing the fluorescence in these dendrites to be isolated from all other filled structures. Here we describe the details of this approach (also called the “periscope” method) and give examples of the kinds of recordings that can be made with this approach.
In Vivo Population Imaging of Dendritic Integration in Neocortex. https://doi.org/10.1007/978-1-62703-785-3_7.
2013
Voltage-dependent calcium channels (VDCCs) serve a wide range of physiological functions and their activity is modulated by different neurotransmitter systems. GABAergic inhibition of VDCCs in neurons has an important impact in controlling transmitter release, neuronal plasticity, gene expression and neuronal excitability. We investigated the molecular signalling mechanisms by which GABA(B) receptors inhibit calcium-mediated electrogenesis (Ca(2+) spikes) in the distal apical dendrite of cortical layer 5 pyramidal neurons. Ca(2+) spikes are the basis of coincidence detection and signal amplification of distal tuft synaptic inputs characteristic for the computational function of cortical pyramidal neurons. By combining dendritic whole-cell recordings with two-photon fluorescence Ca(2+) imaging we found that all subtypes of VDCCs were present in the Ca(2+) spike initiation zone, but that they contribute differently to the initiation and sustaining of dendritic Ca(2+) spikes. Particularly, Ca(v)1 VDCCs are the most abundant VDCC present in this dendritic compartment and they generated the sustained plateau potential characteristic for the Ca(2+) spike. Activation of GABA(B) receptors specifically inhibited Ca(v)1 channels. This inhibition of L-type Ca(2+) currents was transiently relieved by strong depolarization but did not depend on protein kinase activity. Therefore, our findings suggest a novel membrane-delimited interaction of the G(i/o)-βγ-subunit with Ca(v)1 channels identifying this mechanism as the general pathway of GABA(B) receptor-mediated inhibition of VDCCs. Furthermore, the characterization of the contribution of the different VDCCs to the generation of the Ca(2+) spike provides new insights into the molecular mechanism of dendritic computation.
Inhibition of dendritic Ca2+ spikes by GABAB receptors in cortical pyramidal neurons is mediated by a direct Gi/o-β-subunit interaction with Cav1 channels. https://doi.org/10.1113/jphysiol.2012.245464.
Dendrites are the main recipients of synaptic inputs and are important sites that determine neurons' input-output functions. This review focuses on thin neocortical dendrites, which receive the vast majority of synaptic inputs in cortex but also have specialized electrogenic properties. We present a simplified working-model biophysical scheme of pyramidal neurons that attempts to capture the essence of their dendritic function, including the ability to behave under plausible conditions as dynamic computational subunits. We emphasize the electrogenic capabilities of NMDA receptors (NMDARs) because these transmitter-gated channels seem to provide the major nonlinear depolarizing drive in thin dendrites, even allowing full-blown NMDA spikes. We show how apparent discrepancies in experimental findings can be reconciled and discuss the current status of dendritic spikes in vivo; a dominant NMDAR contribution would indicate that the input-output relations of thin dendrites are dynamically set by network activity and cannot be fully predicted by purely reductionist approaches.
Active Properties of Neocortical Pyramidal Neuron Dendrites. https://doi.org/10.1146/annurev-neuro-062111-150343.
Distinct populations of layer 1 inhibitory neurons inhibit or disinhibit layer 5 pyramidal cells. A massive patch-clamp recording effort, tapping up to eight cells simultaneously, maps their influences on the cortical network.
The yin and yang of cortical layer 1. https://doi.org/10.1038/nn.3317.
A basic feature of intelligent systems such as the cerebral cortex is the ability to freely associate aspects of perceived experience with an internal representation of the world and make predictions about the future. Here, a hypothesis is presented that the extraordinary performance of the cortex derives from an associative mechanism built in at the cellular level to the basic cortical neuronal unit: the pyramidal cell. The mechanism is robustly triggered by coincident input to opposite poles of the neuron, is exquisitely matched to the large- and fine-scale architecture of the cortex, and is tightly controlled by local microcircuits of inhibitory neurons targeting subcellular compartments. This article explores the experimental evidence and the implications for how the cortex operates.
A cellular mechanism for cortical associations: an organizing principle for the cerebral cortex. https://doi.org/10.1016/j.tins.2012.11.006.
Modulation of Distal Calcium Electrogenesis by Neuropeptide Y1 Receptors Inhibits Neocortical Long-Term Depression. https://doi.org/10.1523/JNEUROSCI.5595-12.2013.
Processing of sensory information from both sides of the body requires coordination of sensory input between the two hemispheres. This coordination is achieved by transcallosal (interhemispheric) fibers that course though the upper cortical layers. In a recent study by Palmer et al. (2012), we investigated the role of this interhemispheric input on the dendritic and somatic activity of cortical pyramidal neurons. This study showed that interhemispheric input evokes GABAB-mediated inhibition in the distal dendrites of layer 5 pyramidal neurons, decreasing the action potential output when paired with contralateral sensory stimulation. In contrast, layer 2/3 pyramidal neurons were not inhibited by interhemispheric input, possibly due to transcallosal fibers evoking more excitation in these neurons than layer 5 neurons. These results highlight both the precise nature of the microcircuitry of interhemispheric inhibition and how the balance between excitation and inhibition is different in the different layers of the cortex. Identifying the cellular and molecular elements involved in interhemipsheric inhibition is crucial not only for understanding higher brain function and but also dysfunction in the diseased brain.
Layer-specific regulation of cortical neurons by interhemispheric inhibition. https://doi.org/10.4161/cib.23545.
The Brain Research through Advancing Innovative Neurotechnologies (BRAIN) Initiative has focused scientific attention on the necessary tools to understand the human brain and mind. Here, we outline our collective vision for what we can achieve within a decade with properly targeted efforts and discuss likely technological deliverables and neuroscience progress.
The Challenge of Connecting the Dots in the B.R.A.I.N.. https://doi.org/10.1016/j.neuron.2013.09.008.
2012
Abstract Dendrites are highly branched neuronal structures that form the main synaptic target of other nerve cells. They are studded with small spines that are the contact points for excitatory inputs. Dendrites integrate these excitatory inputs, along with inhibitory inputs, in the form of brief depolarising or hyperpolarizing synaptic potentials, and generate the output of the neuron. This signal is usually in the form of an action potential initiated near the soma. Although early models assumed that the dendrites were passive, we now know that they have many voltage dependent conductances distributed heterogeneously over the surface of the arbours. These conductances lead to various forms of signal modulation and localised regenerative activity that greatly expand the computational complexity of CNS neurons. Key Concepts: Dendrites comprise approximately 95% of the neuronal surface and are the main place where synaptic contacts are made. Dendritic architecture is stereotypical for different neurons types and probably relates to the integrative function of these cells. Synaptic potentials originate in the dendrites and propagate decrementally to the axon hillock where the output sodium spike is usually generated. Dendrites contain voltage‐dependent channels distributed in ways characteristic to each cell type, which modulate the propagation of synaptic potentials. These voltage‐dependent channels can also support regenerative action potentials, which either back‐propagate over the dendrites from the somatic region, or are initiated locally in the dendrites. These dendritic action potentials are complex. They are primarily sodium dependent, calcium dependent, or, in some cases, involve the ligand‐gated NMDA receptor. The function of these dendritic action potentials is not always clear. One possibility is to control the calcium concentration in different parts of the cell, which can then regulate synaptic plasticity or other signalling mechanisms. Another possibility is to influence the integration of synaptic potentials and ultimately determine the output of the neuron.
Dendrites. https://doi.org/10.1002/9780470015902.a0000092.pub3.
The precise timing of events in the brain has consequences for intracellular processes, synaptic plasticity, integration and network behaviour. Pyramidal neurons, the most widespread excitatory neuron of the neocortex have multiple spike initiation zones, which interact via dendritic and somatic spikes actively propagating in all directions within the dendritic tree. For these neurons, therefore, both the location and timing of synaptic inputs are critical. The time window for which the backpropagating action potential can influence dendritic spike generation has been extensively studied in layer 5 neocortical pyramidal neurons of rat somatosensory cortex. Here, we re-examine this coincidence detection window for pyramidal cell types across the rat somatosensory cortex in layers 2/3, 5 and 6. We find that the time-window for optimal interaction is widest and shifted in layer 5 pyramidal neurons relative to cells in layers 6 and 2/3. Inputs arriving at the same time and locations will therefore differentially affect spike-timing dependent processes in the different classes of pyramidal neurons.
The Time Window for Generation of Dendritic Spikes by Coincidence of Action Potentials and EPSPs is Layer Specific in Somatosensory Cortex. https://doi.org/10.1371/journal.pone.0033146.
Observing dendritic activity in active animals presents great challenges and there are currently not many options available. This protocol describes the fiber-optic-based "periscope" method for recording from populations of layer 5 (L5) neocortical pyramidal dendrites. This is an inexpensive, convenient, and robust method for recording dendritic activity during behavior. The limitations of this approach are discussed and tips are provided for how it might be adapted for different purposes and brain areas.
Fiber-optic calcium monitoring of dendritic activity in vivo. https://doi.org/10.1101/pdb.prot067835.
Exposure to alcohol in utero is a well known cause of mental retardation in humans. Using experimental models of fetal alcohol spectrum disorder, it has been demonstrated that cortical pyramidal neurons and their projections are profoundly and permanently impaired. Yet, how the functional features of these cells are modified and how such modifications impact cognitive processes is still unknown. To address this, we studied the intrinsic electrophysiological properties of pyramidal neurons in young adult rats (P30–P60) exposed to ethanol inhalation during the first week of postnatal life (P2–P6). Dual whole-cell recordings from the soma and distal apical dendrites were performed and, following the injection of depolarizing current into the dendrites, layer 5 neurons from ethanol-treated (Et) animals displayed a lower number and a shorter duration of dendritic spikes, attributable to a downregulation of calcium electrogenesis. As a consequence, the mean number of action potentials recorded at the soma after dendritic current injection was also lower in Et animals. No significant differences between Et and controls were observed in the firing pattern elicited in layer 5 neurons by steps of depolarizing somatic current, even though the firing rate was significantly lower in Et animals. The firing pattern and the firing rate of layer 2/3 neurons were not affected by alcohol exposure.
Early Exposure to Alcohol Leads to Permanent Impairment of Dendritic Excitability in Neocortical Pyramidal Neurons. https://doi.org/10.1523/JNEUROSCI.5520-11.2012.
The spatiotemporal control of neuronal excitability is fundamental to the inhibitory process. We now have a wealth of information about the active dendritic properties of cortical neurons including axonally generated sodium action potentials as well as local sodium spikelets generated in the dendrites, calcium plateau spikes, and NMDA spikes. All of these events have been shown to be highly modified by the spatiotemporal pattern of nearby inhibitory input which can drastically change the output firing mode of the neuron. This means that particular populations of interneurons embedded in the neocortical microcircuitry can more precisely control pyramidal cell output than has previously been thought. Furthermore, the output of any given neuron tends to feed back onto inhibitory circuits making the resultant network activity further dependent on inhibition. Network activity is therefore ultimately governed by the subcellular microcircuitry of the cortex and it is impossible to ignore the subcompartmentalization of inhibitory influence at the neuronal level in order to understand its effects at the network level. In this article, we summarize the inhibitory circuits that have been shown so far to act on specific dendritic compartments in vivo.
Inhibitory Regulation of Dendritic Activity in vivo. https://doi.org/10.3389/fncir.2012.00026.
Dendrites carry signals between synapses and the soma and play a central role in neural computation. Although they contain many nonlinear ion channels, their signal-transfer properties are linear under some experimental conditions. In experiments with continuous-time inputs, a resonant linear two-port model has been shown to provide a near-perfect fit to the dendrite-to-soma input-output relationship. In this study, we focused on this linear aspect of signal transfer using impedance functions that replace biophysical channel models in order to describe the electrical properties of the dendritic membrane. The membrane impedance model of dendrites preserves the accuracy of the two-port model with minimal computational complexity. Using this approach, we demonstrate two membrane impedance profiles of dendrites that reproduced the experimentally observed two-port results. These impedance profiles demonstrate that the two-port results are compatible with different computational schemes. In addition, our model highlights how dendritic resonance can minimize the location-dependent attenuation of signals at the resonant frequency. Thus, in this model, dendrites function as linear-resonant filters that carry signals between nonlinear computational units.
A compartmental model of linear resonance and signal transfer in dendrites. https://doi.org/10.1162/NECO_a_00366.
Interhemispheric inhibition is thought to mediate cortical rivalry between the two hemispheres through callosal input. The long-lasting form of this inhibition is believed to operate via g-aminobutyric acid type B (GABAB) receptors, but the process is poorly understood at the cellular level. We found that the firing of layer 5 pyramidal neurons in rat somatosensory cortex due to contralateral sensory stimulation was inhibited for hundreds of milliseconds when paired with ipsilateral stimulation. The inhibition acted directly on apical dendrites via layer 1 interneurons but was silent in the absence of pyramidal cell firing, relying on metabotropic inhibition of active dendritic currents recruited during neuronal activity. The results not only reveal the microcircuitry underlying interhemispheric inhibition but also demonstrate the importance of active dendritic properties for cortical output.
The Cellular Basis of GABA <sub>B</sub> -Mediated Interhemispheric Inhibition. https://doi.org/10.1126/science.1217276.
2010
Fluorescent calcium (Ca(2+)) indicator proteins (FCIPs) are promising tools for functional imaging of cellular activity in living animals. However, they have still not reached their full potential for in vivo imaging of neuronal activity due to limitations in expression levels, dynamic range, and sensitivity for reporting action potentials. Here, we report that viral expression of the ratiometric Ca(2+) sensor yellow cameleon 3.60 (YC3.60) in pyramidal neurons of mouse barrel cortex enables in vivo measurement of neuronal activity with high dynamic range and sensitivity across multiple spatial scales. By combining juxtacellular recordings and two-photon imaging in vitro and in vivo, we demonstrate that YC3.60 can resolve single action potential (AP)-evoked Ca(2+) transients and reliably reports bursts of APs with negligible saturation. Spontaneous and whisker-evoked Ca(2+) transients were detected in individual apical dendrites and somata as well as in local neuronal populations. Moreover, bulk measurements using wide-field imaging or fiber-optics revealed sensory-evoked YC3.60 signals in large areas of the barrel field. Fiber-optic recordings in particular enabled measurements in awake, freely moving mice and revealed complex Ca(2+) dynamics, possibly reflecting different behavior-related brain states. Viral expression of YC3.60 - in combination with various optical techniques - thus opens a multitude of opportunities for functional studies of the neural basis of animal behavior, from dendrites to the levels of local and large-scale neuronal populations.
Optical recording of neuronal activity with a genetically-encoded calcium indicator in anesthetized and freely moving mice. https://doi.org/10.3389/fncir.2010.00009.
Layer 6 (L6) pyramidal neurons are the only neocortical pyramidal cell type whose apical dendrite terminates in layer 4 rather than layer 1. Like layer 5 pyramidal neurons, they participate in a feedback loop with the thalamus and project to other cortical areas. Despite their unique location in the cortical microcircuit, synaptic integration in dendrites of L6 neurons has never been investigated. Given that all other neocortical pyramidal neurons perform active integration of synaptic inputs via local dendritic spike generation, we were interested to establish the apical dendritic properties of L6 pyramidal neurons. We measured active and passive properties of the apical dendrites of L6 pyramidal neurons in the somatosensory region of rat cortical slices using dual patch-clamp recordings from somata and dendrites and calcium imaging. We found that L6 pyramidal neurons share many fundamental dendritic properties with other neocortical pyramidal neurons, including the generation of local dendritic spikes under the control of dendritic inhibition, voltage-dependent support of backpropagating action potentials, timing-dependent dendritic integration, distally located I h channels, frequency-dependent Ca 2+ spike activation, and NMDA spike electrogenesis in the distal apical dendrite. The results suggest that L6 pyramidal neurons integrate synaptic inputs in layer 4 similar to the way other neocortical pyramidal neurons integrate input to layer 1. Thus, L6 pyramidal neurons can perform a similar associational task operating on inputs arriving at the granular and subgranular layers.
Properties of Layer 6 Pyramidal Neuron Apical Dendrites. https://doi.org/10.1523/JNEUROSCI.2254-10.2010.
The mechanisms underlying memory formation in the hippocampal network remain a major unanswered aspect of neuroscience. Although high-frequency activity appears essential for plasticity, salience for memory formation is also provided by activity in ventral tegmental area (VTA) dopamine projections. Here, we report that activation of dopamine D1 receptors in dentate granule cells (DGCs) can preferentially increase dendritic excitability to both high-frequency afferent activity and high-frequency trains of backpropagating action potentials. Using whole-cell patch clamp recordings, calcium imaging, and neuropeptide Y to inhibit postsynaptic calcium influx, we found that activation of dendritic voltage-dependent calcium channels (VDCCs) is essential for dopamine-induced long-term potentiation (LTP), both in rat and human dentate gyrus (DG). Moreover, we demonstrate previously unreported spike-timing-dependent plasticity in the human hippocampus. These results suggest that when dopamine is released in the dentate gyrus with concurrent high-frequency activity there is an increased probability that synapses will be strengthened and reward-associated spatial memories will be formed.
Dopamine modulates synaptic plasticity in dendrites of rat and human dentate granule cells. https://doi.org/10.1073/pnas.1011558107.
2009
Live action role playing: die Entwicklung realer Kompetenzen in virtuellen Welten. https://doi.org/.







